Collective Risk Dilemma
A collective-risk social dilemma arises when a group must cooperate to reach a common target in order to avoid the risk of collective loss while each individual is tempted to free-ride on the contributions of others. In contrast to
the prisoners' dilemma or public goods games, the collective-risk dilemma encompasses the risk that all individuals lose everything. These characteristics have potential relevance for dangerous climate change and other risky social
dilemmas. Climate change relief is truly the most influential cooperative act we can be a part of, especially since our decisions now may influence the decisions of future generations. However, collaborative prospects are disheartening due to the uncertainties associated with Climate change and the diversity of the actors involved (eg. wealth). Cooperation is costly to the individual and it only benefits all individuals if the common target is reached.
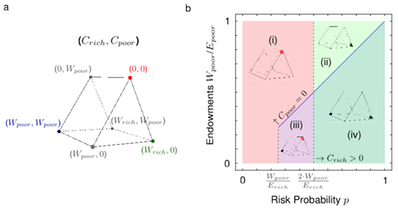
An individual thus invests without guarantee that the investment is worthwhile for anyone. If there are several subsequent stages of investment, it is not clear when individuals should contribute. Thus, in this game, the timing of contributions becomes a strategic variable that allows individuals to interact and influence one another. We investigate the strategic behavior using an evolutionary model of such collective-risk dilemmas in a finite population; every individual takes part in many games and successful behaviors spread in the population. Individual contributions depend on the stage of the game and on the sum of contributions made so far. Thus far prediction have shown that, the best strategy was to `wait and see'.
This was consistent under homogeneity, heterogeneity, target uncertainty, across round number and various group sizes. However one key difference in heterogeneity, the rich players contribute in excess in oder to compensate for the missing contributions of the poor.
Avian Host-Parasite Mafia Behaviour
Mafia like behavior, where individuals cooperate under the threat of punishment, occurs not only in humans, but is also observed in several animal species. Observations suggest that avian hosts tend to accept a certain degree of parasitism in order to avoid retaliating punishment from the brood parasite. To understand under which conditions it will be beneficial for a host to cooperate, we model the interaction between hosts and parasites as an evolutionary game. In our model, the host's behavior is plastic, and thus, its response depends on the previous interactions with the parasite. We find that such learned behavior in turn is crucial for the evolution of retaliating parasites. The abundance of this kind of mafia behavior oscillates in time and does not settle to an equilibrium. Our results suggest that retaliation is a mechanism for the parasite to evade specialisation and to induce acceptance by the host.
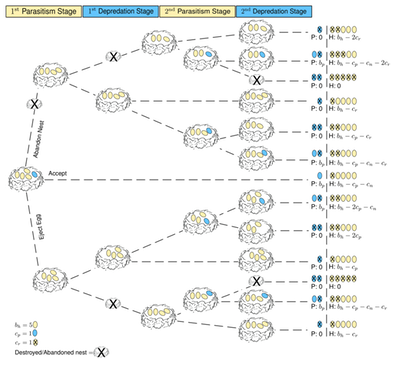
We model the interaction between hosts (e.g. magpies, Pica pica, and prothonotary warblers, Protonotaria citrea) and parasites (e.g. cuckoos, Clamator glandarius, and cowbirds, Molothrus ater) as an evolutionary game. These species tend to interact with each other over several seasons. In each season a parasite can lay more than a single egg in the host’s nest without fear that the parasite nestling will evict its siblings out of the nest; both the young of the host and parasite are raised together 1, 12, 22 . Thus, in this system there are repeated interactions within a breeding season. We classify the host-parasite interactions into four stages: (i) During the first parasitism stage the parasite lays a certain number of eggs in the nest and the host will choose to accept these eggs or to reject them. (ii) If the host rejects, the parasite could depredate and destroy the host’s nest during the first depredation stage. This stage occurs after the host rejects, which could be any time between egg laying and nestling hatching 12. (iii) The second parasitism occurs only when the parasite returns to the same host to lay additional eggs after rejection or after the host reclutches. (iv) The second depredation stage occurs after a host rejects the returning parasite.
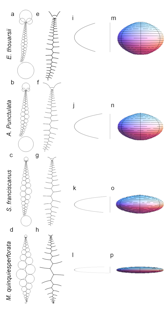
Theoretical Morphology
Echinoids have an endoskeletal system which is ideal for studying
calcified structures such as development of vertebrate skeletons.
However, understanding echinoid skeletal (test) growth has proven
challenging to analyse solely on the basis of any one approach or
process. Therefore, theoretical models have been developed to understand
growth and form of echinoid tests. Herein, Holotestoid, a computational
model of echinoid test growth is described. The model incorporates
mathematical principles (e.g., close-packing), physical principles
(e.g., interface between coalescing bubbles) and biological processes
(e.g., echinoid ontogenic processes). It is the first computational
model that emulates all five ontogenic processes involved in test growth
(plate growth, plate addition, plate interaction, plate gapping, and
visceral growth) using a geometrical representation and three analogies
(coalescing bubble, circle-packing, and catenary chains). The emulated
processes are used to predict plate size, plate shape, and test shape.
Size-Assortative Mating
Size-assortative mating is a nonrandom association of body size between members of mating pairs and is expected to be common in species with mutual preferences for body size. In this study, we investigated whether there is direct evidence for size-assortative mating in two species of pipefishes, Syngnathus floridae and S. typhle, that share the characteristics of male pregnancy, sex-role reversal, and a polygynandrous mating system. We take advantage of microsatellite-based “genetic-capture” techniques to match wild-caught females with female genotypes reconstructed from broods of pregnant males and use these data to explore patterns of size-assortative mating in these species. We also develop a simulation model to explore how positive, negative, and antagonistic preferences of each sex for body size affect size-assortative mating. Contrary to expectations, we were unable to find any evidence of size-assortative mating in either species at different geographic locations or at different sampling times. Furthermore, two traits that potentially confer a fitness advantage in terms of reproductive success, female mating order and number of eggs transferred per female, do not affect pairing patterns in the wild. Results from model simula- tions demonstrate that strong mating preferences are unlikely to explain the observed patterns of mating in the studied populations. Our study shows that individual mating preferences, as ascertained by laboratory-based mating trials, can be decoupled from realized patterns of mating in the wild, and therefore, field studies are also necessary to determine actual patterns of mate choice in nature. We conclude that this disconnect between preferences and assortative mating is likely due to ecological constraints and multiple mating that may limit mate choice in natural populations.
